

发布:2026-05-28 浏览:9 次
 本篇文献分享发表于中科院二区杂志Alzheimer's Research & Therapy。本公众号所发布内容旨在与大家分享学术新知,促进交流学习,版权归原作者或原出处所有,感谢各位学者的辛勤成果。
本篇文献分享发表于中科院二区杂志Alzheimer's Research & Therapy。本公众号所发布内容旨在与大家分享学术新知,促进交流学习,版权归原作者或原出处所有,感谢各位学者的辛勤成果。
1.引言
阿尔茨海默病是老年人痴呆的主要原因,其病理特征为β-淀粉样蛋白斑块和tau蛋白形成的神经原纤维缠结沉积。该疾病的进展遵循从早期阶段至严重痴呆阶段的连续过程,称为阿尔茨海默病谱系。在该谱系中,最早的阶段是主观认知下降,该阶段可能逐渐进展为轻度认知障碍,最终发展为典型的阿尔茨海默病痴呆。
人脑被视为一个由相对稳定结构网络支持的动态功能系统,功能网络的时域波动被认为是复杂认知功能和意识产生的基础。在此框架下,基于静息态功能磁共振成像时间序列的动态功能连接分析在揭示多种神经系统疾病(包括精神分裂症、重性抑郁障碍和阿尔茨海默病)的脑连接模式方面显示出巨大潜力。在动态功能连接方法中,滑动窗方法已在阿尔茨海默病谱系中被广泛采用。既往研究发现,主观认知下降患者默认模式网络的动态功能连接变异性降低,认知控制网络与感觉运动网络之间的连接也表现出减弱。另一项研究则揭示轻度认知障碍患者中央执行网络与背侧注意网络之间的动态功能连接变异性增加。还有研究指出,脑动态状态可揭示轻度认知障碍的不同进展,并可能有助于早期识别高危轻度认知障碍患者。所有这些研究表明,动态功能连接异常在疾病早期即已出现,并可能随临床进展而演变。然而,滑动窗方法受限于窗口长度的选择和对瞬时或细微波动的敏感性有限,这在主观认知下降阶段尤为成问题,因为该阶段的脑功能改变通常较为细微。
相比之下,一种称为主导特征向量动力学分析的新方法已被开发出来。主导特征向量动力学分析捕捉每个时间点血氧水平依赖信号的瞬时相位锁定模式,无需滑动窗口,特别适用于检测短暂或弱表达的网络状态。重要的是,主导特征向量动力学分析在检测疼痛的细微纵向变化、抑郁症复发以及治疗相关改变方面已显示出高敏感性。这些研究突显了主导特征向量动力学分析在追踪阿尔茨海默病谱系疾病进展中的潜力。在阿尔茨海默病和轻度认知障碍中应用主导特征向量动力学分析的初步研究揭示了停留时间和占有率的独特改变,提示存在异常的动态脑状态。然而,将主导特征向量动力学分析应用于主观认知下降人群的研究仍然稀缺,且跨阿尔茨海默病谱系阶段的动态功能连接功能障碍轨迹尚未得到系统表征。
近年来,转录组学和神经影像遗传学的进展使得探索宏观脑功能障碍的分子机制成为可能。将空间基因表达数据与神经影像表型相整合,有望桥接阿尔茨海默病的微观和宏观病理。例如,关于阿尔茨海默病中铁代谢失调的研究揭示,与病理性铁沉积相关的基因富集于蛋白磷酸化和金属离子稳态通路,并主要在小胶质细胞和谷氨酸能神经元中表达。此外,有研究报道阿尔茨海默病患者海马和内侧颞叶的结构改变与调控化学突触传递的基因表达密切相关。利用Juelich细胞构筑图谱,另一项研究进一步揭示了遗传变异(特别是与胆碱能通路相关的变异)与阿尔茨海默病谱系中动态功能连接改变相关。随着疾病进展,越来越多的证据表明阿尔茨海默病谱系个体的全局脑不稳定性逐渐下降。尽管如此,动态功能连接改变的转录组学基础以及这些改变如何随疾病连续过程演变仍不清楚。
在此研究中,旨在利用主导特征向量动力学分析探究阿尔茨海默病谱系中阶段依赖的动态功能连接改变,并进一步阐明其遗传和神经生物学基础。假设脑网络动态随认知下降在阿尔茨海默病谱系中呈渐进性恶化,且这些脑网络功能障碍与阿尔茨海默病病理相关的基因表达模式存在空间关联。通过整合脑影像与转录组数据,此研究旨在:(1)表征主观认知下降、轻度认知障碍和阿尔茨海默病阶段的独特动态脑状态;(2)识别与认知下降相关的动态功能连接模式;(3)揭示脑功能不稳定性背后的转录组学特征。此研究结果将为阿尔茨海默病谱系早期脑网络功能障碍提供机制性见解,并支持开发用于早期检测和疾病监测的遗传信息神经影像生物标志物。
2.材料与方法
2.1参与者
此研究收集了人口学数据,包括年龄、性别、教育程度、高血压和糖尿病状况及神经系统疾病史。所有受试者均由记忆专科医生进行标准化神经心理学评估,包括简易精神状态检查、画钟测验以及连线测验A部分和B部分。
参与者分为四组:健康对照(69人)、主观认知下降(83人)、轻度认知障碍(56人)和阿尔茨海默病(31人)。健康对照组无认知障碍或临床异常。主观认知下降受试者符合既定的SCD-plus临床框架;轻度认知障碍病例按标准诊断;阿尔茨海默病的诊断符合2011年美国国家衰老研究所-阿尔茨海默病协会指南中“临床诊断阿尔茨海默病”的标准。排除标准包括:(1)其他神经退行性痴呆(如血管性痴呆、路易体痴呆和额颞叶痴呆);(2)药物或精神障碍引起的认知障碍;(3)符合《精神障碍诊断与统计手册第五版》的严重精神障碍(如重性抑郁症、精神分裂症)。根据《赫尔辛基宣言》,此研究经华中科技大学同济医学院同济医院伦理委员会批准。所有参与者均提供知情同意书。
2.2磁共振成像采集
磁共振成像扫描使用3.0T GE Discovery 750 MRI系统进行。受试者呈仰卧位,扫描范围从头顶至颅底。
静息态功能磁共振成像采用梯度回波平面成像序列采集。主要参数包括:重复时间/回波时间=2000/30 ms,矩阵大小64×64,视野220×220 mm,层厚3 mm,翻转角90°,共36层连续轴位层,采集185个时间点。结构磁共振成像采用3D-T1 BRAIN Volume序列,参数:重复时间/回波时间=7.1/2.7 ms,视野256×256 mm,采集矩阵256×256,激励次数=1,层厚=1 mm。
2.3磁共振数据处理
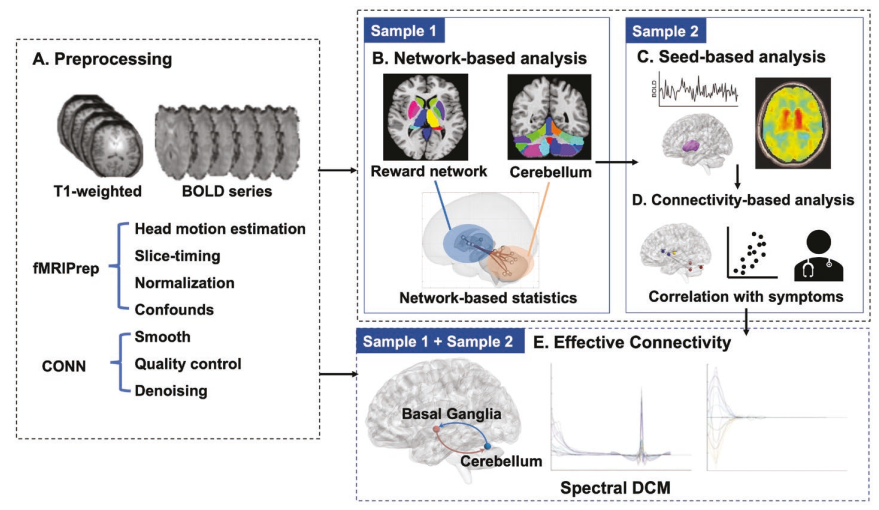
此研究的数据处理流程如图1所示。主要处理步骤包括功能磁共振成像预处理、主导特征向量动力学分析及其与临床评分的相关性,以及将显示组间差异的瞬时脑状态与基因表达及相关生物学过程相关联的富集分析。
图1分析流程示意图。
2.3.1功能磁共振数据预处理
使用CONN工具箱进行功能磁共振成像数据预处理。丢弃前5个时间点以使扫描仪信号稳定,保留180个时间点用于分析。通过调整每个时间点的运动参数进行头动估计和校正,随后进行时间层校正。具体而言,采用12个运动参数进行头动回归,包括3个平移参数和3个旋转参数及其一阶导数。为减轻运动相关伪影的影响,剔除异常时间点,标记运动过度的异常时间点。运动排除标准定义为:帧间位移阈值0.9 mm,全局BOLD信号变化阈值5个标准差。将功能数据配准至对应的3D-T1 BRAIN结构图像,所有图像空间归一化至MNI空间,并进行灰质、白质和脑脊液的组织分割。最后,采用6 mm半高全宽高斯核进行空间平滑。预处理后,在CONN工具箱中通过回归白质和脑脊液信号的前五个主成分进行去噪处理。采用Schaefer100-Yeo7图谱将大脑划分为若干脑区,再从每个脑区中提取时间序列。
2.3.2主导特征向量动力学分析与脑状态特征
对于动态连接分析,使用以往研究中的主导特征向量动力学分析代码,并在自动化处理阶段额外去除5个时间点。使用二阶巴特沃斯滤波器对BOLD时间序列进行带通滤波(0.02-0.10 Hz)以减轻生理噪声。随后通过希尔伯特变换计算每个感兴趣区的瞬时相位信号,实现基于相位的连接分析。随后采用余弦距离法计算每对脑区在每个重复时间点的相位差,得到动态相位锁定矩阵,量化脑区间的相位对齐。通过计算该矩阵的主特征向量,以显示所有脑区BOLD相位的主要方向特征。使用K-means对特征向量进行聚类。聚类数K为5-10,涵盖静息态功能磁共振成像文献中报道的典型功能网络数量。通过最小Davies-Bouldin指数选择最佳K值(最终K=10)。
脑状态动态特征采用三个关键指标表征:(1)占有率,量化在每个状态中花费的总时间比例;(2)停留时间,测量连续访问某状态的平均持续时间;(3)状态转换概率,反映状态间切换的可能性。使用一般线性模型评估这些动态功能连接指标的组间差异,控制年龄、性别和受教育年限,统计显著性采用Bonferroni校正。对于10个状态的占有率和停留时间比较,显著性阈值设为p < 0.005(0.05/10)。对于涉及100个有向转换的转换概率分析,阈值设为p < 5×10⁻⁴(0.05/100)。结果部分报告的p值为Bonferroni校正后的值,通过将原始p值乘以检验次数获得。
2.3.3脑状态特征与认知相关性
为探究脑功能状态指标与认知表现之间的关系,在四个参与者组中采用一般线性模型。调整年龄、性别和受教育年限作为协变量,检验每个定义脑状态的停留时间和占有率与认知功能评估指标之间的相关性,包括简易精神状态检查、画钟测验、连线测验A部分和B部分。
2.4基因类别富集分析
对于表现出显著组间差异的状态,进行基因类别富集分析(GCEA)以探究其潜在分子机制。首先,使用abagen工具箱将艾伦人脑图谱的区域微阵列基因表达数据映射至Schaefer100-Yeo7脑区。然后使用ABAnnotate工具箱将空间脑状态与源自艾伦人脑图谱数据的转录组特征相关联。该工具箱在多个基因转录类别中进行富集分析:(1)基因本体论富集分析,聚焦于生物学过程类别,来自DAVID数据库;(2)发育性脑区基因表达谱,使用BrainSpan数据库的RNA-seq数据表征发育性脑区的基因表达;(3)神经元细胞类型关联分析,基于PsychENCODE数据库的神经元细胞类型标志物。
为降低空间自相关的影响,并避免因脑区共用皮层结构带来的分析偏倚,本研究采用计算类别内共表达模式的方法,而不是直接进行基因水平的相关分析。具体而言,计算表型效应与基因表达之间的Spearman相关性。然后通过取z变换相关性的加权平均值(按类别特异性表达水平缩放)将这些相关性聚合为表型类别评分。通过10,000次自旋置换生成零分布评估显著性,随后进行错误发现率多重比较校正。
2.5统计分析
使用SPSS 12.0软件分析人口学数据。采用Shapiro-Wilk检验评估正态性,Levene检验评估方差齐性。由于连续变量(年龄、教育年限、简易精神状态检查、画钟测验、连线测验A部分、连线测验B部分)不满足正态性假设,对所有连续变量采用Kruskal-Wallis检验,而分类变量(性别、高血压、糖尿病)采用卡方检验进行比较。
在脑海科技云平台中,内置了基于主导特征向量动力学分析、神经影像与基因关联分析模块,支持用户批量处理数据,并确保每一步参数设置都有据可查。平台提供完整的操作文档、培训视频及更新日志体系,便于用户快速掌握。此外,平台的项目管理模块可清晰记录数据筛选标准、排除被试原因、分析版本等信息,极大提升了研究的透明度和可复现性。读者可对照思考如何在平台中实现类似的分析流程。感兴趣可联系预约产品演示。
3.结果
3.1人口学特征
参与者的分组及人口学特征如下:健康对照组69人,主观认知下降组83人,轻度认知障碍组56人,阿尔茨海默病组31人。四组年龄中位数分别为64岁、66岁、64.5岁和66岁,教育年限中位数分别为12年、15年、12年和12年。性别构成方面,女性/男性比例分别为34:35、59:24、29:27和12:19。四组在年龄、高血压和糖尿病方面无显著差异(p > 0.05)。然而,性别分布(p = 0.004)和教育程度(p < 0.001)存在显著统计学差异。所有神经心理学评估(包括简易精神状态检查、画钟测验、连线测验A部分和B部分)均显示出显著的组间差异(均p < 0.001)。
3.2阿尔茨海默病谱系中瞬时脑状态的差异
采用主导特征向量动力学分析框架内的K-means聚类分析,此研究识别出十个全脑瞬时功能状态。其中一个状态以全脑广泛的相位一致性为特征,称为全脑同步状态(状态10)。其余九个状态表现出显著的区域特异性同步模式。(1)双侧半球控制网络与默认模式网络的协调激活(状态1,Cont-DMN);(2)双侧半球背侧注意网络与感觉运动网络的高度同步(状态2,DAN-SMN);(3)双侧半球视觉网络与感觉运动网络的高度同步(状态3,Vis-SMN);(4)双侧半球视觉网络与控制网络的强同步(状态4,Vis-Cont);(5)双侧半球突显/腹侧注意网络与控制网络及默认模式网络的高度同步(状态5,SVAN-Cont-DMN);(6)双侧半球背侧注意网络与边缘网络的高度同步(状态6,DAN-LIM);(7)双侧半球视觉网络与默认模式网络的强同步(状态7,Vis-DMN);(8)双侧半球感觉运动网络、背侧注意网络与默认模式网络的高度同步(状态8,SMN-DAN-DMN);(9)左侧半球边缘网络与默认模式网络的高度同步(状态9,LIM-DMN)。
与健康对照相比,轻度认知障碍患者在状态7(Vis-DMN)的占有率显著降低(Pbfn*10=0.030,β=0.03)(图2A)。相比之下,阿尔茨海默病患者在状态4(Vis-Cont)、状态5(SVAN-Cont-DMN)和状态6(DAN-LIM)的停留时间(状态4:Pbfn*10=0.025,β=1.12;状态5:Pbfn*10=0.013,β=1.08;状态6:Pbfn*10=0.013,β=1.16)和占有率(状态4:Pbfn*10=0.006,β=0.04;状态5:Pbfn*10=0.026,β=0.04;状态6:Pbfn*10=0.013,β=0.03)均显著降低。此外,阿尔茨海默病患者在状态10(Global)的停留时间(Pbfn*10=0.001,β=-3.18)和占有率(Pbfn*10<0.001,β=-0.13)均显著增加。阿尔茨海默病患者在状态2(DAN-SMN)的占有率也显著升高(Pbfn*10=0.028,β=-0.05),在状态3(Vis-SMN)的停留时间显著降低(Pbfn*10=0.010,β=1.05)(图2B)。值得注意的是,健康对照与主观认知下降组之间在任何脑状态的停留时间或占有率上均未观察到显著差异。
在脑状态转换方面,与健康对照相比,主观认知下降患者从状态8(SMN-DAN-DMN)向状态10(Global)转换的概率显著升高(Pbfn*100=0.010,β=-0.033)。然而,主观认知下降患者从状态4(Vis-Cont)向状态3(Vis-SMN)转换的概率(Pbfn*100=0.025,β=0.024)、从状态5(SVAN-Cont-DMN)向状态1(Cont-DMN)转换的概率(Pbfn*100=0.022,β=0.038),以及从状态7(Vis-DMN)向状态1(Cont-DMN)转换的概率(Pbfn*100=0.049,β=0.035)均显著降低(图2C)。
与健康对照相比,轻度认知障碍患者从状态10(Global)向状态9(LIM-DMN)转换的概率增加(Pbfn*100=0.023,β=-0.033),从状态7(Vis-DMN)向状态2(DAN-SMN)转换的概率也增加(Pbfn*100=0.048,β=-0.037)。然而,轻度认知障碍患者从状态2(DAN-SMN)向状态3(Vis-SMN)转换的概率(Pbfn*100=0.033,β=0.057)、从状态6(DAN-LIM)向状态9(LIM-DMN)转换的概率(Pbfn*100=0.036,β=0.035),以及从状态10(Global)向状态5(SVAN-Cont-DMN)转换的概率(Pbfn*100=0.007,β=0.064)均降低(图2D)。
在阿尔茨海默病患者中,与健康对照相比,多个状态转换存在显著差异。阿尔茨海默病患者从状态1(Cont-DMN)向状态10(Global)转换的概率(Pbfn*100=0.018,β=-0.030)、从状态2(DAN-SMN)向状态10(Global)转换的概率(Pbfn*100=0.037,β=-0.047)、从状态3(Vis-SMN)向状态10(Global)转换的概率(Pbfn*100=0.002,β=-0.104)、从状态4(Vis-Cont)向状态10(Global)转换的概率(Pbfn*100=0.003,β=-0.089)、从状态5(SVAN-Cont-DMN)向状态10(Global)转换的概率(Pbfn*100=0.009,β=-0.090),以及停留在状态10(Global)的概率(Pbfn*100=0.014,β=-0.150)均显著升高。此外,阿尔茨海默病患者从状态3(Vis-SMN)向状态2(DAN-SMN)转换的概率也更高(Pbfn*100=0.019,β=-0.044)。然而,阿尔茨海默病患者从其他多个状态转换的概率较低,特别是停留在状态3(Vis-SMN)的概率(Pbfn*100=0.003,β=0.123)、停留在状态4(Vis-Cont)的概率(Pbfn*100=0.012,β=0.113)、停留在状态5(SVAN-Cont-DMN)的概率(Pbfn*100=0.020,β=0.115)、停留在状态6(DAN-LIM)的概率(Pbfn*100=0.008,β=0.130),以及从状态1(Cont-DMN)向状态4(Vis-Cont)转换的概率(Pbfn*100=0.047,β=0.039)、从状态2(DAN-SMN)向状态3(Vis-SMN)转换的概率(Pbfn*100=0.027,β=0.079)、从状态6(DAN-LIM)向状态1(Cont-DMN)转换的概率(Pbfn*100=0.047,β=0.023)、从状态9(LIM-DMN)向状态4(Vis-Cont)转换的概率(Pbfn*100=0.045,β=0.026)均较低(图2E)。
图2采用GLM对十种脑状态间的停留时间、占据率及转移概率进行组间比较。
3.3与神经心理学评估相关的脑状态特征
十个脑状态与神经心理学评估结果的相关性分析显示,状态3(Vis-SMN)的停留时间(p = 0.006,β = 0.182)和状态5(SVAN-Cont-DMN)的停留时间(p = 0.011,β = 0.167)与简易精神状态检查得分呈显著正相关,而状态10(Global)的停留时间与简易精神状态检查得分呈显著负相关(p = 0.002,β = -0.202)。此外,状态3(Vis-SMN)和状态6(DAN-LIM)的停留时间与连线测验A部分得分呈显著负相关(状态3,Vis-SMN:p = 0.015,β = -0.188;状态6,DAN-LIM:p = 0.002,β = -0.234),而状态10(Global)的停留时间与连线测验A部分得分呈显著正相关(p < 0.001,β = 0.339)(图3A)。
状态3(Vis-SMN)、状态4(Vis-Cont)、状态5(SVAN-Cont-DMN)和状态6(DAN-LIM)的占有率与简易精神状态检查得分呈显著正相关(状态3,Vis-SMN:p = 0.047,β = 0.131;状态4,Vis-Cont:p = 0.016,β = 0.159;状态5,SVAN-Cont-DMN:p = 0.041,β = 0.137;状态6,DAN-LIM:p = 0.029,β = 0.143),而状态2(DAN-SMN)和状态10(Global)的占有率与简易精神状态检查得分呈显著负相关(状态2,DAN-SMN:p = 0.003,β = -0.200;状态10,Global:p < 0.001,β = -0.232)。此外,状态6(DAN-LIM)的占有率与连线测验A部分得分呈显著负相关(p = 0.009,β = -0.198),而状态10(Global)的占有率与连线测验A部分得分呈显著正相关(p < 0.001,β = 0.360)(图3B)。然而,未发现十个脑状态的特征与画钟测验或连线测验B部分得分之间存在显著相关。
图3与神经心理学评估相关的3种脑状态特征。
3.4与瞬时脑状态相关的BrainSpan发育基因表达
对十个脑状态相关基因的时空表达模式进行了探究。结果发现,与状态5(SVAN-Cont-DMN)相关的基因在婴儿期、青春期和成年期的皮层区域表现出显著富集的时空表达谱(图4A)。具体而言,在婴儿期,这些基因在眶额叶皮层(OFC;PFDR = 0.048)、内侧前额叶皮层(MFC;PFDR = 0.048)、背外侧前额叶皮层(DFC;PFDR = 0.048)、初级躯体感觉皮层(S1C;PFDR = 0.048)和顶下皮层(IPC;PFDR = 0.043)显著富集(图4B)。在青春期和成年期,这些基因在眶额叶皮层(青春期:PFDR = 0.032;成年期:PFDR = 0.048)、内侧前额叶皮层(青春期:PFDR = 0.048;成年期:PFDR = 0.032)和颞下回(ITG)(青春期:PFDR = 0.048;成年期:PFDR = 0.032)特异性富集(图4C)。相比之下,其余9个状态相关的基因在大脑中未表现出统计学显著的空间聚类模式。
图4 BrainSpan发育期基因表达谱分析与阿尔茨海默病谱系中短暂脑状态的关联性研究。
3.5与瞬时脑状态相关的神经元细胞类型
对十个脑状态相关的基因集进行细胞类型富集分析。结果显示,与状态5相关的基因集(SVAN -ContDMN)在兴奋性神经元亚型中呈现显著富集,包括Ex4(PFDR =0.007)、Ex5(PFDR =0.007)和Ex7(PFDR =0.026)。该基因集还在抑制性神经元亚型In1中显示出特异性表达信号(PFDR =0.047)(图5)。解剖学上,Ex4、Ex5和Ex7代表具有不同层状分布的皮层投射神经元:Ex4主要分布于第5层,Ex5/Ex7则主要位于第6层。In1是一种GABA能抑制性中间神经元,特别是源自内侧神经节隆起(MGE)的快速放电亚型,且呈小清蛋白阳性(PV⁺)特征。相比之下,与其他9种状态相关的基因集在兴奋性或抑制性神经元亚型中均未显示出统计学显著富集。
图5与阿尔茨海默病谱系中短暂脑状态相关的神经元细胞类型。
3.6与瞬时脑状态相关的基因本体论分析
对十个脑状态进行的基因本体论分析显示,仅状态5在离子通道活性相关的分子功能方面表现出特异性基因富集。该状态中富集最强的基因包括GLRA2、GLRB和GLRA3,主要基因本体论术语为“细胞外甘氨酸门控离子通道活性”(GO:0016933;PFDR = 0.0006)及其亚类“细胞外甘氨酸门控氯离子通道活性”(GO:0016934;PFDR = 0.0006),两者均参与调节抑制性神经递质传递。(图6).值得注意的是,在细胞组分或生物过程类别中均未观察到显著富集现象(PFDR >0.05)。此外,在其余9种状态中,所有GO功能类别(分子功能、细胞组分或生物过程)均未出现具有统计学意义的信号。
图6与阿尔茨海默病谱系中短暂脑状态相关的基因本体论富集分析。
4.讨论
参考文献



